情報通信研究機構(NICT) 未来ICT研究所
神戸フロンティア研究センター 神経網ICT研究室 行動神経生物学プロジェクト(山元研究室)
私たちの研究室では、動物の行動が起こる仕組みを、分子(遺伝子)・細胞(脳)・個体のレ ベルで解明していくことを目指しています。
![]()
研究内容
7.幹細胞
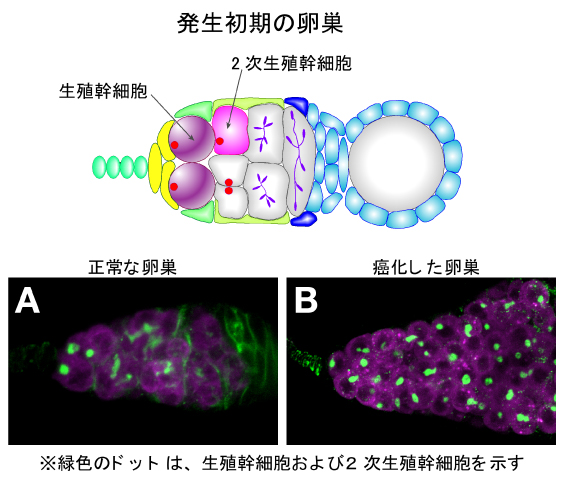
人工幹細胞、iPSは、山中伸弥教授のノーベル賞受賞で日本津々浦々、知らぬ者なき存在となった。体内の幹細胞は何度も分裂して娘細胞を生み出し、その娘細胞の多くは分裂することなく分化して個体の各部で決まった働きの担い手となる。一方、娘細胞の片割れはもとの幹細胞の性質を受け継いで、次の分裂に向かう。つまり幹細胞は自己保存的な分裂によって自分自身を生み出しつつ、絶えず成熟へと向かう“普通の細胞”をも作り出している。なればこそ、いろいろな臓器のもととなる細胞たちをiPSによって作ることが可能なのである。しかし、この“自己保存”と“普通の細胞”の産み分けを誤ると、細胞の新生ができなくなったり、逆に制御のきかない増殖による腫瘍の形成に直結する。この絶妙なバランスはいったいどのようにして保たれているのだろうか。そのコントロールは、幹細胞自身に組み込まれたプログラムによるところも大きいが、それに劣らず、幹細胞の“ゆりかご”として働く周囲の細胞たちの寄与があって初めて可能である。この“ゆりかご”はニッチと呼ばれている。ニッチと幹細胞の二人三脚で進む増殖と分化のバランスのとれたコントロール、その理解を可能にするもっともすぐれたシステムがキイロショウジョウバエの卵巣にある。ニッチを構成する細胞と生殖幹細胞を体内で全て同定できる稀有なシステムがここにあるのだ。我々は、性行動異常の突然変異体、fickleの解析過程で、この変異体の卵巣に腫瘍が形成されることに気付いた44)。この腫瘍が形成される仕組みを研究していくうちに、fickle変異体ではニッチの細胞でトランスポゾンの転移が頻繁に起こることを発見した。トランスポゾンは利己的遺伝子の代表格である。トランスポゾンの転移がおこるとDNAに傷がつき、精子や卵でこれが起こるとそれは直ちに突然変異となって現れる。それを抑えているのはPiwiと呼ばれるタンパク質である。Piwiタンパク質は、piRNAという短いRNA(マイクロRNA:miRNA)をたくさん身にまとい(結合し)、トランスポゾンに張り付いてその動きを止める働きをしている。piRNAの配列にはトランスポゾンの配列と相同性があり、そのためゲノムに入り込んだ利己的遺伝子であるトランスポゾンの配列を見つけ出してそれに結合するのである45)。piRNAはトランスポゾンの宿主(この場合、キイロショウジョウバエ)のゲノムに存在する遺伝子群(たとえばflamencoという遺伝子座)から転写されてできる。ということは、このゲノムの遺伝子群のDNA配列がトランスポゾンと相同性があることになる。つまり、このゲノム上の遺伝子群は、もとをただせばよそ者のトランスポゾンそのものだということである。結局、よそ者のトランスポゾンを使ってより最近に侵入したトランスポゾンをやっつける、“毒を持って毒を制する”戦略を生物は使っているということだ。そして、今や昨日の敵は今日の友、ディフェンスのためのなくてはならない飛び道具になっているわけである。piRNAの一部は、宿主のゲノム中でまっとうなタンパク質をコードしている月並みな遺伝子のmRNAの3’UTR(非翻訳領域)からも作られてくるというから驚きである。きっとこの3’UTRの配列も、もとをただせばトランスポゾンだったのだろう。fickle変異の原因遺伝子産物は、この驚くべき宿主―利己的遺伝子のせめぎあいの真っただ中で生殖幹細胞を守る先兵として活躍していたのだ。その機構の解明は、まだまだ序に着いたばかりである。
Btk29A promotes Wnt4 signaling in the niche to terminate germ cell proliferation in Drosophila.
Science 343, 294-297.
PIWI-interacting small RNAs: the vanguard of genome
defence.
Nat. Rev. Mol. Cell
Biol. 12, 246-258.
Designed by CSS.Design Sample