私たちは、以下に示す主要なシナプスタンパク質遺伝子の突然変異体を解析することによって各タンパク質の機能を知り、どのようにしてシナプスでの伝達が行われるかを調べてきた。孵化できない致死突然変異体も解析可能であるという利点をもつショウジョウバエ胚を材料として、神経筋シナプスでシナプス電流の計測を行っている(図2)。

図2. ショウジョウバエ胚神経筋シナプスで発生するシナプス電流の記録
そして、シナプトタグミン1がシナプス伝達を活動電位に同期していること(図3)を最初に報告した。この発見は、後にAugustine lab, Chapman lab, Sudhof labなどによって、哺乳類においてもその普遍性が確認された。さらに、シナプトブレビンが活動電位に誘起されるシナプス伝達に必須であるということについても、やはりハエを用いた我々の遺伝学的な解析が突破口となり、後にSudhof labの哺乳類を使った研究で確認され、シナプス伝達の普遍的な分子細胞メカニズムの解明に貢献した。現在もシナプトタグミン7の機能などに関して、MITのLittleton教授と共同研究を続けている。
n-シナプトブレビン
Yoshihara et al., (1999) J. Neurosci., 19: 2432-2441.
Yoshihara et al., (2000) J. Neurosci., 20: 8315-8322.
シナプトタグミン1
Yoshihara and Littleton (2002) Neuron 36: 897-908
Yoshihara et al., (2010) Proc. Natl. Acad. Sci. USA 107: 14869-14874
シナプトタグミン7(未発表)
N-タイプカルシウムチャネル(未発表)
グルタミン酸受容体
Yoshihara et al., (2005) Science 310:858-863
総説は
Yoshihara et al., (2002) Current Opinions in Neurobiology 13: 315-323.
Yoshihara and Montana (2004)
The Neuroscientist 10: 566-574.
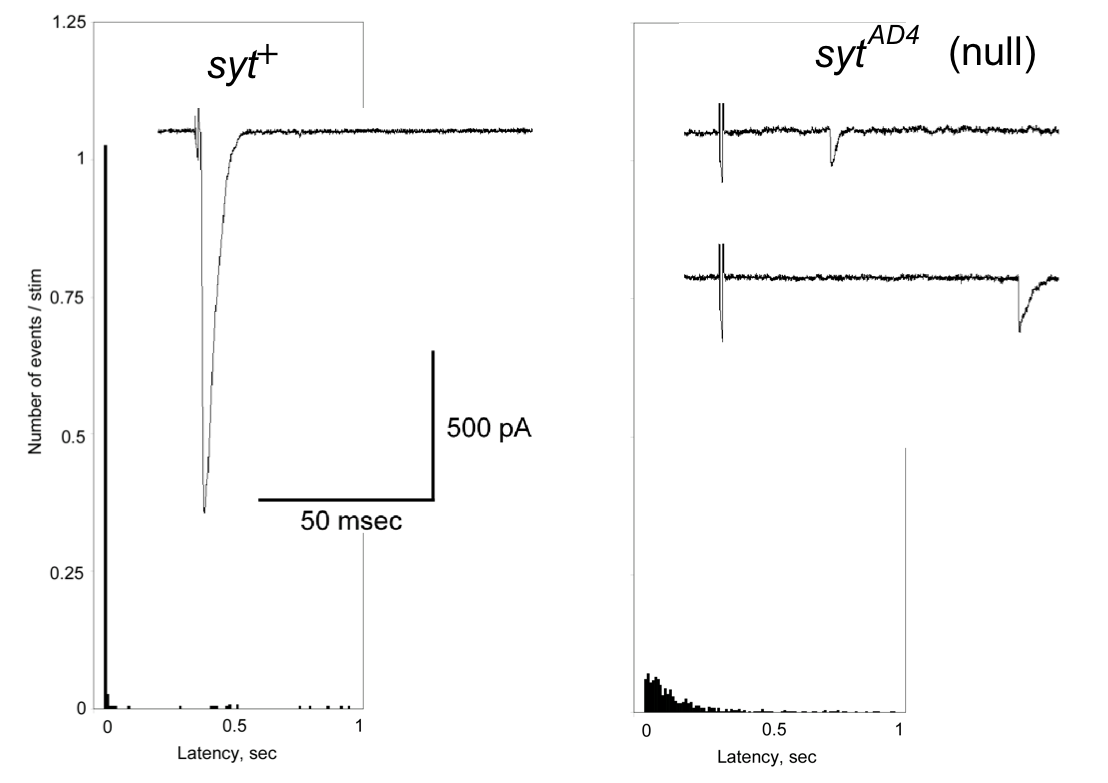
図3. ショウジョウバエ野生型(左)とSyt
1欠損突然変異体(右)の胚神経筋シナプス伝達。グラフは横軸を神経刺激からシナプス伝達の遅れにとったヒストグラム。 Syt 1のない突然変異体(右)では、野生型(左)よりシナプス伝達が遅くなる。Syt 1 がシナプス小胞のすばやい放出を可能にしているためである。
(2)シナプスの発達へ進む
(3)シナプスの可塑性 (ショウジョウバエ胚シナプスの生理学)へ移動する
(4)フィーディング・ニューロン上のシナプス変化として”記憶形成を見る”へ移動する
研究内容TOPへ